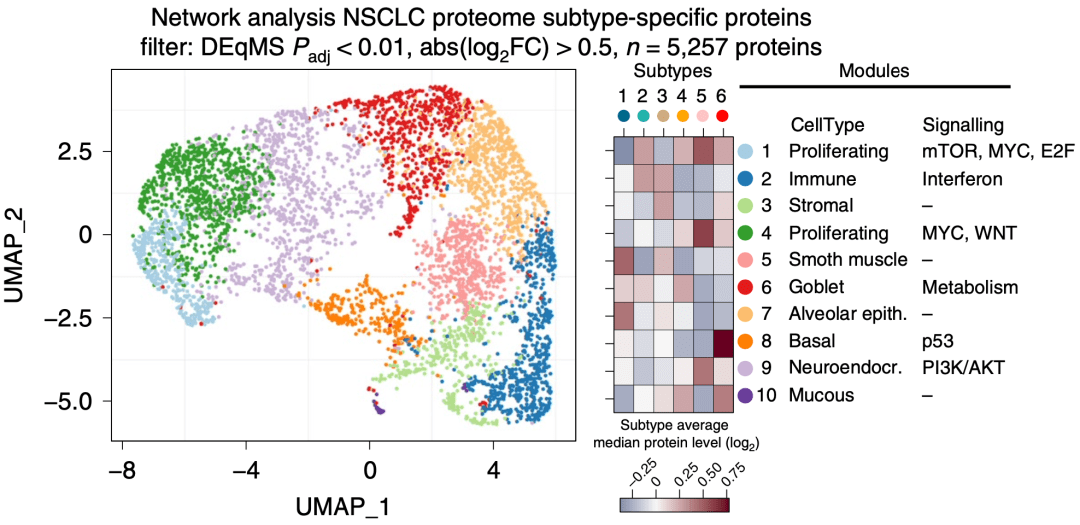
以上 突变分布模式与蛋白网络分析结果是一致的:亚型4富集STK11突变蛋白;亚型5富集E2F1/MYC信号通路及RB1突变蛋白;亚型6含有p53信号通路和TP53突变蛋白 。 亚型5中全部SqCC样本都具有RB1突变 , 唯一没有聚类到亚型5中的LCNEC样本 , 具有STK11和KEAP1突变 。 这说明该 蛋白组分析有效地反映了NSCLC的不同组织学分类各自具有的主要癌症分子特征和驱动性信号通路变化 。
文章图片
▲ 突变分布模式与蛋白网络分析结果是一致的
为了探究蛋白组信息与已知致癌基因之间更具体的关系 , Lehti?团队选定了951种“癌症驱动相关蛋白”(CDRPs) 。
在测定的832种CDRPs中 , 有291种在至少一个样本中显示出异常值 (此处异常值设定为蛋白水平高或低于所有样本中值的三倍) 。 85%的样本具有至少一种致癌基因表达异常值 , 26%样本则至少含有5个致癌基因表达异常值 。
所有6个亚型中 , 亚型5有最高的平均过表达致癌基因数 , 且通常包括转录激活因子MYB 。 在AC为主的亚型1–4中 , 亚型4是平均过表达致癌基因数量最多的 , 并大都包括受体酪氨酸激酶RET 。 更进一步的分析显示 , NSCLC中过表达的除了广为人知的驱动基因 (例如EGFR , ERBB2和KRAS) , 还包括一些不常被提到的其他致癌基因 , 例如致癌性激酶SGK1 。
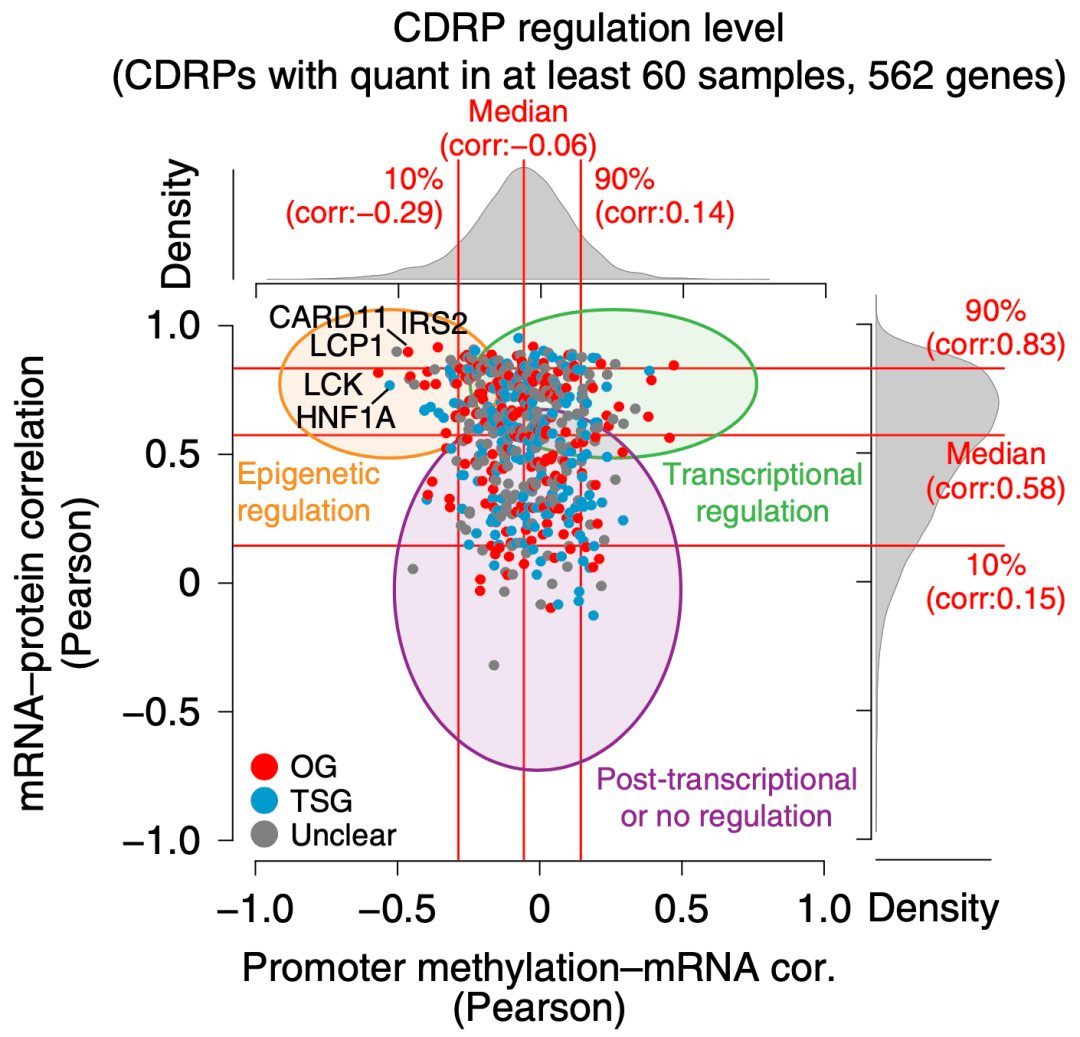
蛋白质谱数据加上基因组范围内的转录组及甲基化组数据 , 大大方便了Lehti?团队分析基因调控水平 。 虽然从整体来看 ,mRNA和蛋白质水平的正相关性很高 , 但仍有一部分CDRP的mRNA水平无法从蛋白组数据解释 , 这些基因的表达很可能受到了表观调控、转录调控或者转录后调控 。
文章图片
▲ 横坐标:启动子甲基化程度 – mRNA水平相关性;纵坐标:mRNA-蛋白质水平相关性 。 黄色圈中包含的CDRPs(例如LCK, HNF1A, LCP1, CARD11和IRS2)有可能是受表观调控的
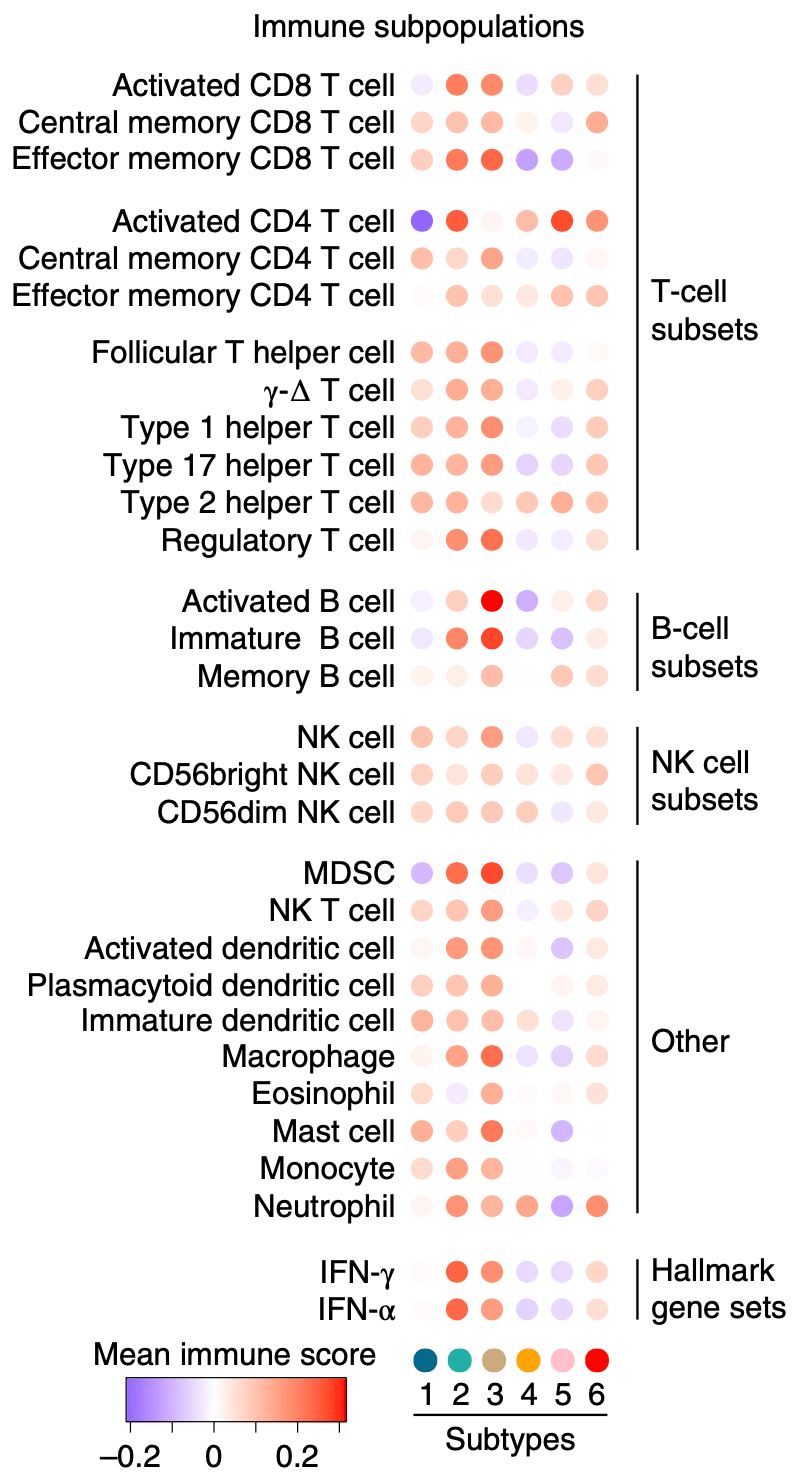
接下来 , Lehti?团队开始分析不同样本中渗透免疫细胞的类型 。 根据Charoentong等人[3]划分的免疫特征 , 他们发现 亚型2中有大量T细胞和干扰素(IFN)信号 , 亚型3中存在很高的B细胞特征信号 , 两者具有已明显分化的免疫响应 。 同时 , 亚型4中所有类型的免疫细胞信号都非常低 , 属于免疫冷型 。
文章图片
▲ 在6个NSCLC蛋白组亚型中渗透免疫细胞的主要类型各不相同 。 MDSC:髓系衍生抑制细胞;NK:自然杀伤细胞
早期研究发现 , 富集B细胞的三级淋巴结构(TLSs)预示着良好的预后[4]和免疫治疗应答[5] 。 对此 , Lehti?团队选取并分析了AC样本中PD-L1高表达(亚型2)或B细胞生物标志物高表达(亚型3)的肿瘤切片 , 验证了亚型3中不仅存在TLSs , 还具有差异显著的组织生长模式 。
那么这种区别会影响新抗原依赖性免疫细胞激活吗?
Lehti?团队对抗原呈递元件(APM)和肿瘤突变负荷(TMB)的分析显示:亚型2具有高APM和高TMB , 亚型3具有高APM和低TMB , 亚型4则具有高TMB和低APM 。 高APM和高TMB意味着新抗原的产生和呈递 , 从而诱导强烈的免疫激活 。 与此同时 , PD-L1是亚型2特征性表达的标记蛋白之一 。 这都表明 ,靶向PD-L1/PD-1免疫检查点的治疗策略 , 对属于亚型2的病例将更为有效 。
接下来 , Lehti?团队进行了免疫检查点的系统研究 。 他们发现 , 抑制性受体(IRs)的蛋白水平总体上与T细胞浸润相关 , 然而IR配体(通常由癌细胞和APCs产生)表达特征非常多样化 , 说明 不同亚型中存在的免疫逃逸机制并不相同 。 最引人注目的是亚型2中高表达的IR配体PD-L1 , 同时大家还注意到另外两种特异性IR配体 , 即亚型4中的FGL1和亚型6中的B7-H4 。
- 卫生室|《乡村医生诊疗口袋书(先行本)》面世
- 解读|《浙江省医院门诊管理办法》政策解读
- 廉东波|《健谈》观点丨浅表性胃炎三五年复查一次 萎缩性胃炎一年一查
- 家长|杭州《青少年皮肤健康状况调研》发布,近半青少年受皮肤疾病困扰
- 公报,监测,国民,体质|一图读懂《第五次国民体质监测公报》
- 作者:鲍秀兰专家团队本文转载自“儿科医生鲍秀兰”(xiehebaoxiulan)来源:金...|孩子老喊腿疼,是长高的预兆吗?如果有这些症状别大意
- 感冒|《气血运行影响心脏功能——心悸篇》
- 幽门螺杆菌感染|《健谈》观点丨突然消瘦、胃痛、黑便,应及时做胃镜检查
- 数据|《胃肠病学》:减肥防早发肠癌,要从20岁开始!科学家发现,20、30岁和约45岁时肥胖,与结直肠癌发病风险上升88-156%有关
- 医学|科普连载:《氢气医学人群试验》(漫画版)孙学军教授 序言